2021年12月22日,国际生物学杂志《PLoS Biology》在线刊出了北京师范大学认知神经科学与学习国家重点实验室邢大军研究组的题为 “Multiple gamma rhythms carry distinct spatial frequency information in primary visual cortex”的文章。该项研究针对视皮层中Gamma振荡的视觉编码规律以及视觉皮层的不同环路连接模式如何产生和调制Gamma振荡的问题,通过多通道电极在多脑区联合记录(视觉皮层V1和外侧膝状体LGN)结合神经解剖及数学建模等手段,首次发现V1中存在三种不同的窄带Gamma振荡,它们分别携带不同的空间频率信息(图1),且有完全不同的神经起源机制。

图1 多个gamma振荡携带了不同的空间频率信息
大脑中的Gamma振荡(25-100Hz)是一种常见于许多脑区的同步神经活动, 它被认为是大脑网络的一种功能性特征,与诸如注意、记忆、学习等正常认知功能紧密相关,在大脑信息处理(如视觉皮层)中也起着重要作用;异常的Gamma振荡常被发现与精神疾病有关。因此对Gamma振荡的功能和环路机制的理解,是解读大脑信息整合机制中的一个前沿问题。尽管Gamma被认为与视觉认知功能有关,但它所表征的视觉信息和产生的神经环路机制仍然不清楚或存在争议。
目前关于Gamma振荡的一个重要问题是:它到底表征什么样的视觉信息?早期研究发现,光条或物体轮廓1–5这类高空间频率的刺激,可以使初级视觉皮层(V1)不同位置的神经元产生强烈Gamma同步振荡(图2A);众多学者由此认为,Gamma振荡可以整合(binding)视觉空间上不同位置的物体轮廓信息,从而表征视觉物体(或者象征整个物体的轮廓)的整体信息。但是最近的一些研究表明,V1对物体表面的神经反应有更强烈的Gamma同步振荡(图2B和C)6–8;Fries等学者据此认为,Gamma振荡表征视觉刺激的表面信息8。最新研究还发现,某些颜色的物体表面可以诱发比其它颜色表面更强的Gamma 7,8。Gamma表征轮廓信息和物体表面信息的矛盾发现,令人费解(图1.3),也挑战了关于Gamma振荡在信息绑定和信息表征功能的现有理论。
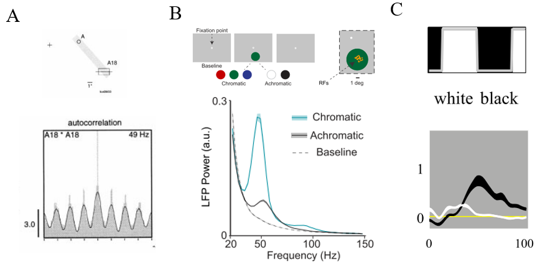
图2 表面或物体刺激引起的Gamma振荡
注:A图展示的是条形刺激在猫的18区中引起强烈的Gamma振荡(频率在49 Hz)。 B图显示了有色(绿松石色),无色(黑色)和基线(灰色)条件下的平均LFP功率谱。 C图显示了V1对4Ca中黑白刺激的平均LFP功率谱。(A图来源于Castelo-Branco et al (1998),B图来源于 Peter et al (2019),C图来源于 Xing et al (2014))
为了解释这个分歧,论文作者对不同空间频率诱发Gamma振荡的情况,以及信息如何通过Gamma进行神经活动编码进行探索,图3A显示了不同空间频率(SFs)移动光栅诱发的典型视觉相关LFP波形。对于大多数记录位点,移动光栅可在不同的空间频率范围内诱发强烈的Gamma振荡(图3B)。令人惊讶的是,不同空间频率的视觉刺激可以在V1中同时诱发三个窄带Gamma振荡。论文作者通过量化发现在V1中存在的这三种不同的窄带Gamma振荡分别处理不同的空间频率(SF)信息,低频Gamma(LG,25-40hz)倾向于高空间频率,中频Gamma(MG,40-65 Hz)和高频Gamma(HG,65-90 Hz)倾向于中低空间频率。
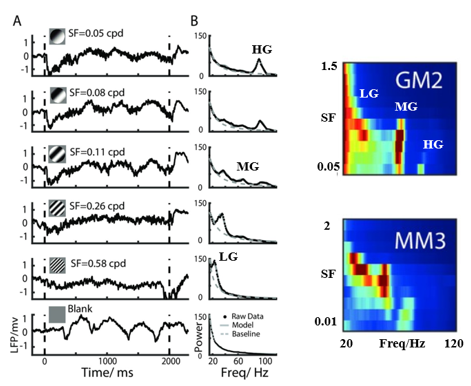
图3 在不同空间频率下诱发出现的三个不同的Gamma振荡
注:A显示了局部场电位(LFP)在不同空间频率(SF)下对漂移光栅刺激的单次试验。黑色虚线表示视觉刺激的开始和偏移。B中的黑点表示LFP在不同空间频率的功率谱,拟合曲线以灰色线绘制。灰色虚线显示描述模型拟合的基线。右图为两只动物(GM2和MM3)功率谱的2维原始数据。横轴为振荡频率,纵轴为不同的空间频率,颜色表示功率。
该文章作者进一步继续测量了视觉刺激表面和边缘引起的Gamma振荡。在屏幕的不同位置显示一个大的黑色或白色正方形(大小为25度,背景为灰色)2秒钟,记录位点的感受野(RF)要么在正方形的边缘上,要么在其表面上(图4左图)。作者发现,当记录位点的感受野位于边缘时,出现低频Gamma振荡(LG);而当记录位点的感受野位于表面时,出现中频Gamma振荡(MG)和高频Gamma振荡(HG)(图4右图)

图4 V1中三个Gamma振荡的表面/边缘偏好
注:左图显示了视觉刺激的演示:相对于感受野的不同位置(表面或边缘)的大而均匀的黑色刺激。右图显示了示例记录位点中表面和边缘条件下的功率谱(边缘条件为紫色曲线,表面条件为蓝色曲线)。
目前Gamma振荡被认为产生于兴奋性和抑制性神经元之间的相互作用9,10。然而,不同的环路和脑区都具有兴奋抑制的相互作用,以及产生Gamma振荡的能力10。一部分前人的工作表明视觉皮层中的Gamma振荡来自于皮层环路之前的视网膜或者外膝体11,12,而另外一部分结果表明,视皮层的Gamma振荡产生于皮层环路(尤其是输出层13)而非皮下核团14。一个自然的问题便是这三个不同的Gamma振荡成分是否有不同的神经起源机制。该项研究进一步使用不同的电极同时记录V1(Utah)和外侧膝状体(LGN,U-probe)(图5A)。图5B显示了两个脑区同时记录的典型示例(来自两只动物)的结果:在V1有三个Gamma振荡成分(图5B,顶行)的时候, LGN仅有一个高频的Gamma振荡成分(HG)(图5B,中行,图5)。LGN的spike和V1的LFP之间的在高频Gamma频段很强的相干性进一步表明,V1的HG来源于皮下LGN(图5B,底行),,而另外两个Gamma振荡产生于视觉皮层。
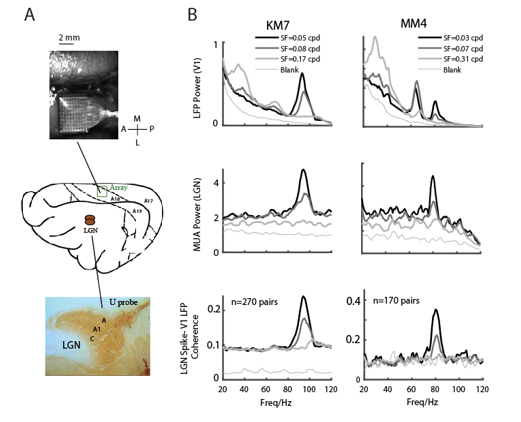
图5 多个Gamma振荡的不同神经起源机制
注:A显示了阵列(V1)和线性阵列(LGN)同时记录的不同大脑区域。B的前两行显示了V1中的LFP功率谱和两只猫同时在不同SF中的MUA功率谱LGN。下一行显示LGN中MUA和V1中LFP之间的相干性。
综上,这项研究针对Gamma相关的两个重要分歧和争议(空间频率的反应特性与不同Gamma振荡的神经环路起源),给出了明确的实验解释;同时,此项研究的实验结果,还验证了邢大军课题组在的另外一篇计算模型15中的预测。这些结果表明了,Gamma振荡主要反映了神经环路的动力学特性;而这些神经环路处理着不同的空间频率信息,同时也是不同空间尺度上神经连接的特征。以上发现对完善Gamma振荡的认知功能理论,深入理解神经信息编码和整合计算原理,解析认知功能与脑环路结构的关系,都具有重要的指导意义;还将为类脑计算的优化,提供实验和理论依据;也对预测和诊断精神疾病以及评估脑功能有着重要意义。

图6 基于三个Gamma振荡的反应特性和神经环路产生机制主要发现的示意图
注:该图说明了基于三次Gamma振荡的反应特性和产生机制。左侧显示的是一幅原始图像(青椒蔬菜),这个图像可以通过视觉系统从皮下核团到皮层区域,在右侧分解为三个不同的信号通道(低、中、高空间频率)。三种颜色(红色、绿色和蓝色)的信号表示三种Gamma。
这项工作得到了国家自然科学基金资助(31371110, 32100831),中央高校基本科研业务费专项资金资助,111项目基金(BP0719032),以及认知神经科学与学习国家重点实验室开放研究基金的资助。本篇文章第一作者为韩传亮博士,通讯作者为邢大军教授。国立台湾大学的叶俊毅教授、上海复旦大学的俞洪波教授以及邢大军课题组其他成员也对此项工作做出了重要贡献。
相关论文链接:
参考文献:
1. Gray, C. M. & Singer, W. Stimulus-specific neuronal oscillations in orientation columns of cat visual cortex. Proc. Natl. Acad. Sci. U. S. A. (1989).
2. Uhlhaas, P. J. & Singer, W. Neural Synchrony in Brain Disorders: Relevance for Cognitive Dysfunctions and Pathophysiology. Neuron (2006).
3. Uhlhaas, P. J. & Singer, W. Abnormal neural oscillations and synchrony in schizophrenia. Nature Reviews Neuroscience (2010).
4. Uhlhaas, P. J. & Singer, W. Neuronal Dynamics and Neuropsychiatric Disorders: Toward a Translational Paradigm for Dysfunctional Large-Scale Networks. Neuron 75, 963–980 (2012).
5. Castelo-Branco, M., Neuenschwander, S. & Singer, W. Synchronization of visual responses between the cortex, lateral geniculate nucleus, and retina in the anesthetized cat. J. Neurosci. 18, 6395–6410 (1998).
6. Gail, A., Brinksmeyer, H. J. & Eckhorn, R. Contour decouples gamma activity across texture representation in monkey striate cortex. Cereb. Cortex 10, 840–850 (2000).
7. Xing, D., Yeh, C. I., Gordon, J. & Shapley, R. M. Cortical brightness adaptation when darkness and brightness produce different dynamical states in the visual cortex. Proc. Natl. Acad. Sci. U. S. A. 111, 1210–1215 (2014).
8. Peter, A. et al. Surface color and predictability determine contextual modulation of V1 firing and gamma oscillations. Elife 8, 1–38 (2019).
9. Xing, D. et al. Stochastic generation of gamma-band activity in primary visual cortex of awake and anesthetized monkeys. J. Neurosci. 32, 13873–13880 (2012).
10. Jia, X., Xing, D. & Kohn, A. No consistent relationship between gamma power and peak frequency in macaque primary visual cortex. J. Neurosci. 33, 17–25 (2013).
11. Saleem, A. B. et al. Subcortical Source and Modulation of the Narrowband Gamma Oscillation in Mouse Visual Cortex. Neuron 93, 315–322 (2017).
12. Storchi, R. et al. Modulation of Fast Narrowband Oscillations in the Mouse Retina and dLGN According to Background Light Intensity. Neuron 93, 299–307 (2017).
13. Xing, D., Yeh, C. I., Burns, S. & Shapley, R. M. Laminar analysis of visually evoked activity in the primary visual cortex. Proc. Natl. Acad. Sci. U. S. A. 109, 13871–13876 (2012).
14. Bastos, A. M., Briggs, F., Alitto, H. J., Mangun, G. R. & Martin Usrey, W. Simultaneous recordings from the primary visual cortex and lateral geniculate nucleus reveal rhythmic interactions and a cortical source for gamma-band oscillations. J. Neurosci. 34, 7639–7644 (2014).
15. Han, C. et al. The Generation and Modulation of Distinct Gamma Oscillations with Local, Horizontal, and Feedback Connections in the Primary Visual Cortex: A Model Study on Large-Scale Networks. Neural Plast. 2021, 8874516 (2021).

