Now, more than ever, increasing exposure to psychosocial stress has become an unavoidable part of our contemporary society as the pace of life rapidly accelerates. Moderate stress can lead to beneficial forms of learning that promote adaptation, whereas stress overload is a key risk factor that elicits the increasing incidence of various psychiatric diseases such as anxiety and depression. Under long-term stress, some people can always overcome difficulties by the continuously improvement of self-regulation and coping skills. Others instead exhibit the exact opposite patterns of impaired performance and even further presage mental disorders. Which factor could reconcile such discrepancy? Trait anxiety, a stable disposition to interpret a wide range of environmental events in a negative way, has been recognized as a vulnerable factor which could account for the seemingly paradoxical effects of stress. However, how trait anxiety and long-term stress interact to affect dynamic decisions during n-back task remains unclear. Is long-term stress a good thing or bad thing for us?
Recently, Prof. Shaozheng Qin’s team of IDG/ McGovern Institute of Brain Science and State Key Laboratory of Cognitive Neuroscience and Learning, Beijing Normal University, and the Prof. Jianhui Wu’s team of Shenzhen University, published their latest empirical study in Cerebral Cortex. They address the questions proposed above by leveraging fMRI and computational modeling of trial-by-trial decision responses to investigate how long-term stress and trait anxiety interact to affect dynamic decision computations during n-back task. In the long-term stress group, participants were recruited from those who have been preparing for the upcoming competitive Chinese National Postgraduate Entrance Exam (CNPEE) for at least 6 months. Only male participants are included to mitigate potential confounds related to menstrual cycles. As expected, individuals in the long-term stress group experienced higher psychological distress than controls, indicating the effectiveness of the natural long-term stress paradigm. Moreover, individuals with higher trait anxiety experienced more psychological distress in general, even after controlling for state anxiety (Fig. 1). These results are in line with previous findings on sustained distress and other symptoms in chronic stress (MacLeod C and R Hagan 1992), which agrees with the psychological view of trait anxiety as a vulnerable phenotype of stress-related psychopathology (Bishop S and S Forster 2013; Weger M and C Sandi 2018).
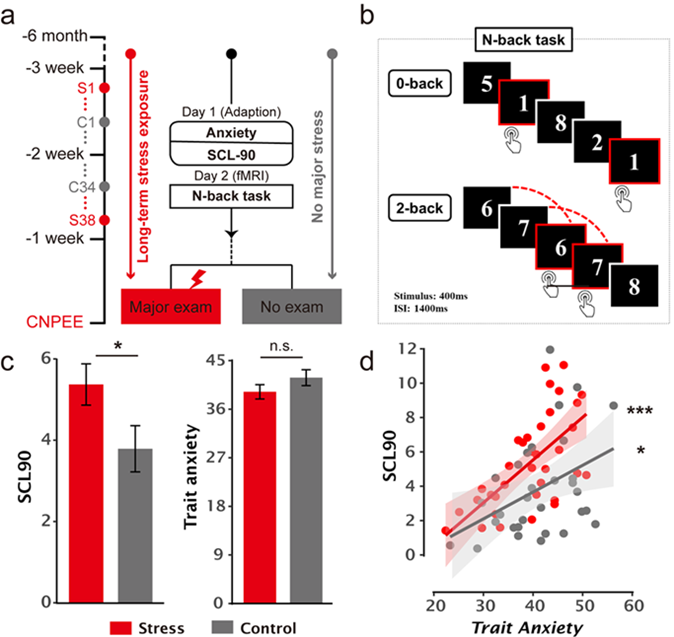
Fig1. Experimental design and the effects of trait anxiety on psychological distress measurements amongst long-term stress and control groups.
Research in sequential-sampling theory posits that the N-back task, analogous to speeded decision-making, can be modeled as an evidence accumulation process during which effective information (namely evidence) extracted from a stream of inherently noisy observations are rapidly accumulated until sufficient evidence reaches the threshold to make a decision and the choice response is then executed(Ratcliff R 1978). For the n-back task, such evidence accumulation process can be considered as accumulating effective information in mind on the exact position of each item in order to make a precise decision whether the current item appeared two positions back in the sequence. A Bayesian hierarchical version of the DDM (HDDM) was implemented to decompose participant’s choice responses in the n-back task into latent decision-making dynamics modulated by free parameters. Of these parameters, the speed of evidence accumulation refers to ‘drift rate’ reflecting the ability to extract effective information from perceived inputs (Ratcliff R and G McKoon 2008). Behaviorally, individuals under long-term stress exhibited faster RTs and drift rate but comparable accuracy than those in controls (Fig 2). In accordance with integrative models of stress, anxiety, and cognitive performance (Derakshan N and MW Eysenck 2009; Edwards EJ et al. 2015), our results show that sustained exposure to exam stress may not impair performance effectiveness (i.e., comparable accuracy), and may enhance processing efficiency (i.e., faster RTs and drift rate) under high cognitive demanding task. However, such enhanced efficiency differs from previously reported cognitive deficits of chronic stress (Arnsten AFT 2009; Lupien SJ et al. 2009). The factor of the interplay of long-term stress with trait anxiety is critical to reconcile this discrepancy (Ebner K and N Singewald 2017; Weger M and C Sandi 2018). When taking individual’s trait anxiety into account, lower trait-anxious individuals exhibited relatively faster drift rate in those under long-term stress than in controls, while accuracy remained at a comparable level across groups and anxiety levels (Fig 3). In other words, stress-induced faster drift rate is driven by low-trait anxious individuals, suggesting that a beneficial form of adaptation to sustained exam stress is likely driven by enhanced processing efficiency in those individuals. This is consistent with previous studies on stress vulnerability reporting that some individuals seem to act as “resilient” agents who can develop adaptive strategies to cope with stress (Franklin TB et al. 2012; Ebner K and N Singewald 2017; Weger M and C Sandi 2018). Furthermore, our observations on high trait-anxious individuals may reflect the recruitment of compensatory strategies to prevent shortfalls in accuracy according to an influential model of cognitive trait anxiety and performance (Eysenck MW et al. 2007).
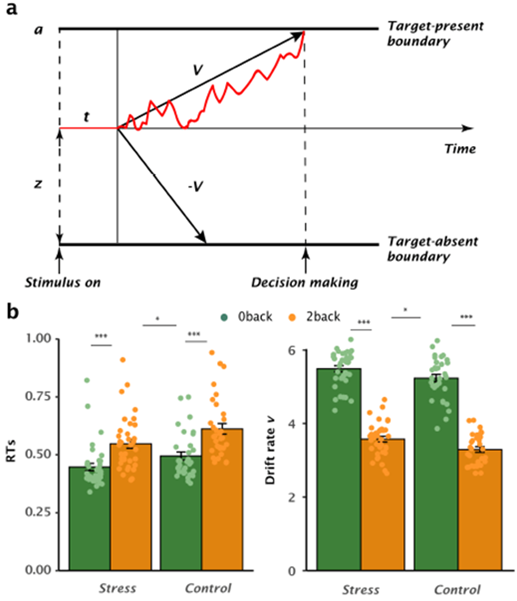
Fig 2. Effects of long-term stress on latent dynamic decision-making during n-back task.
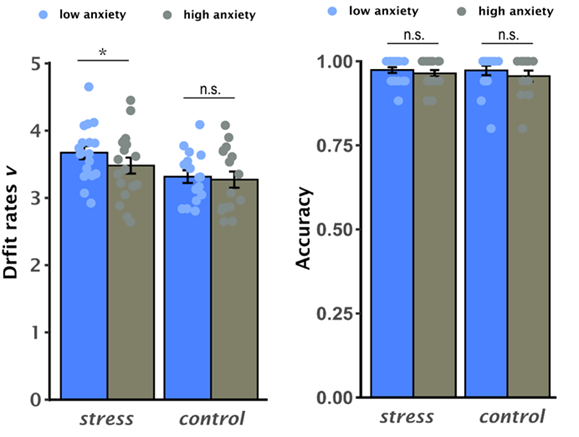
Fig 3. Averaged latent drift rate (left panel) and accuracy (right panel) in the 2-back condition when splitting participants into low and high trait anxiety in both stress and control groups.
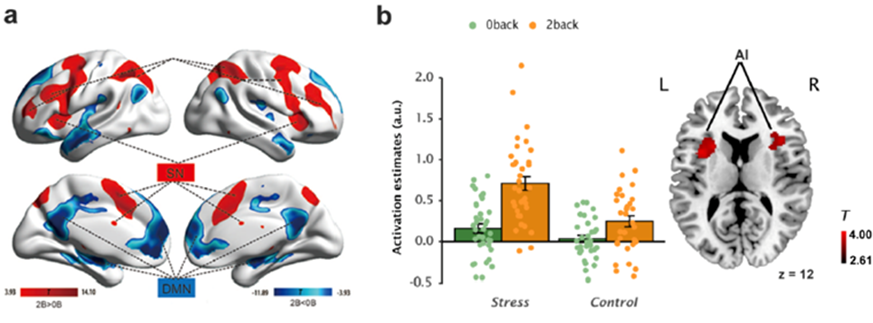
Fig 4. Brain regions showing main effects of WM load and long-term stress.
At the brain activation level, consistent with previous study (Phillips ML et al. 2008), individuals under long-term stress exhibited a general hyper-activation in the anterior insula regardless of WM load (Fig 4). Moreover, long-term stress led to greater WM-related activation in core nodes of the SN including the anterior insula and dACC, but less WM-related deactivation in the DMN regions (Fig 5). The anterior insula and dACC are thought to support salience processing, emotional awareness of distress, and executive function (Menon V ; Menon V and LQ Uddin 2010).
Based on the dual competition model of emotion-cognition interaction (Pessoa L 2009), such SN regions are responsible for reallocating neural resources to resolve competition between emotional processing and executive function. Thus, greater SN engagement under long-term stress may reflect recruitment of additional effort to cope with stress reactivity and to regulate stress-induced distress feelings along with related thoughts that are irrelevant to the WM task. Stress-induced task-irrelevant internal thoughts were indicated by accompanying less DMN deactivation, which parallels the empirical findings of aberrant DMN suppression in psychiatric diseases such as depression(Whitfield-Gabrieli S and JM Ford 2012).
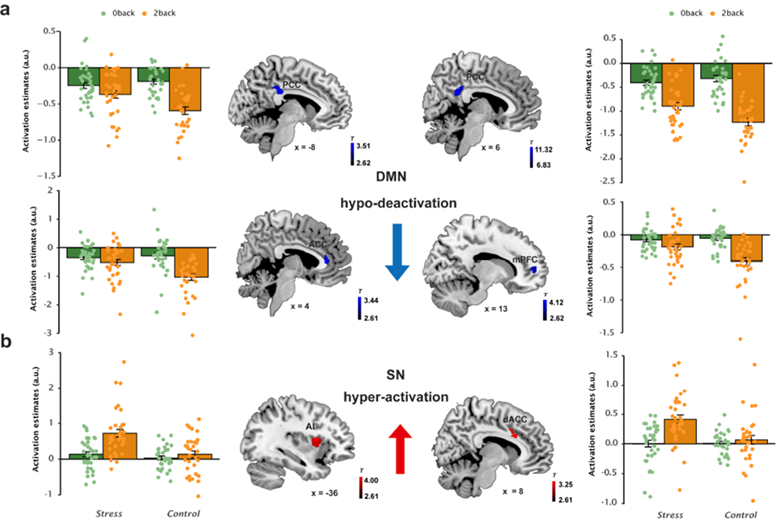
Fig 5. Long-term stress shifts the balance between WM-related brain activation and deactivation.
Separate multi-group SEMs were conducted to test the mediating effects of executive task-related activity (i.e., IPS and MFG) on the association between trait anxiety and drift rate in long-term stress and control groups. Individuals with higher trait anxiety under long-term stress exhibited an indirect association with slower drift rate through higher frontoparietal activity but not those in controls under high task demand (Fig 6). The lack of a mediation effect in the control group suggests that the FPN serves as a mediator only in individuals under long-term stress. Such mediation effects parallel the cognitive models of anxiety and related studies showing that high trait anxiety impairs processing efficiency but not performance effectiveness on tasks involving executive function under stressful conditions (Edwards EJ et al. 2015). As discussed earlier, anxious individuals may recruit additional resources as a compensatory strategy to achieve comparable performance effectiveness (Calvo MG et al. 1994; Calvo MG 1996; Eysenck MW et al. 2007). Given the dlPFC and IPS have been associated with drift rate in the process of evidence accumulation in human and non-human primate studies with single cell recording(Sereno M et al. 2001; Roitman JD and MN Shadlen 2002; Liu T and TJ Pleskac 2011), our observed higher frontoparietal engagement provides neuroimaging evidence to suggest that higher trait-anxious individuals under long-term stress tend to recruit more neural resources to maintain comparable task performance at the cost of the speed of evidence accumulation to make correct decisions when task demand increases.
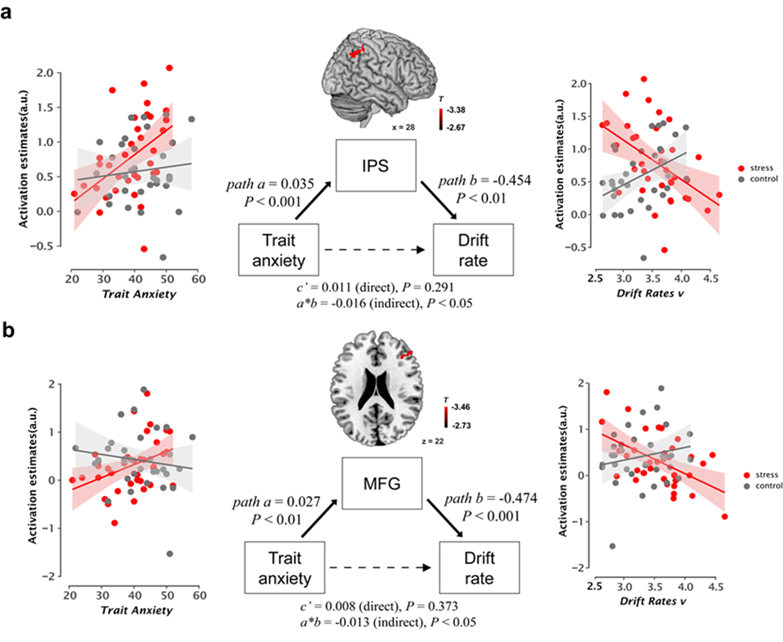
Fig 6. The relation between trait anxiety, brain activity, and drift rate.
At the brain network level, we found greater FPN-DMN decoupling during WM under long-term stress than controls regardless of WM load (Fig 7). Stronger FPN-DMN decoupling here may reflect recruitment of additional effort to suppress task-irrelevant internal thoughts, while performing the goal-directed task (Hampson M et al. 2010). This notion is also in line with our observed faster RTs and drift rate under long-term stress. Critically, under high task demand, individuals with higher trait anxiety exhibited stronger decoupling between DMN and FPN regions in those under long-term stress but not in controls, and such stronger decoupling was then associated with slower drift rate (Fig 7). This again provides evidence to suggest that high trait-anxious individuals might recruit additional resources relying on FPN-DMN decoupling, along with the elevated FPN activity mentioned above, to make correct responses and prevent a shortfall in accuracy at the cost of processing efficiency when task goal is difficult to achieve.
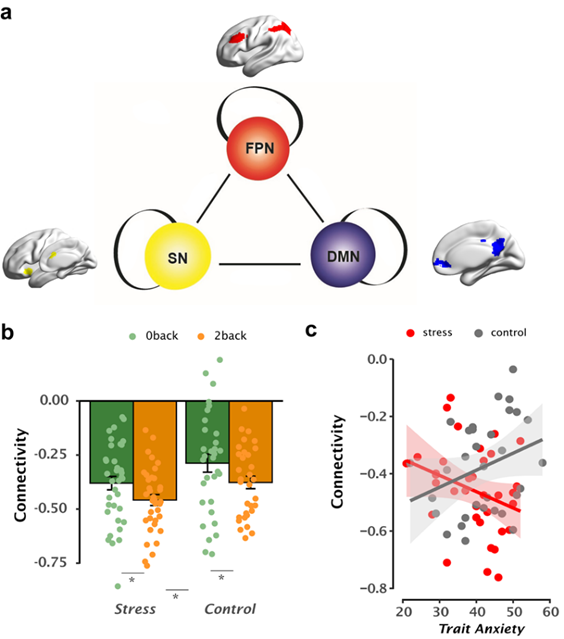
Fig 7. Inter-network connectivity modulated by trait anxiety under long-term stress.
In conclusion, our results provide new insights into the interaction of psychological stress and cognition: Long-term stress can trigger low-trait anxious individual’s potential to perform tasks better, and high trait anxiety itself dose not necessarily lead to cognitive deficits. Instead, high trait anxiety works in tandem with long-term stress to maintain comparable task performance at the expense of the speed of evidence accumulation by recruiting more neural resources. Therefore, no matter what type of person you are, let’s learn to make friends with stress in our life!
This study is the extension and continuation of our research work in the field of human emotional cognition and psychological stress in recent 5 years (Zhuang et al, Nat Hum Behav 2021; Xiong et al. Prog Neurobiol 2021; Jiang et al., Biol Psychiatry 2021; Tian et al., Biol Psychiatry 2021; Liu et al., Nat Commun 2016). This work was supported by the National Natural Science Foundation of China (32130045, 31522028, 81571056, 82021004, 2014NT15, 31771246) and the Fundamental Research Funds for the Central Universities. Liu Liangying, a master student of Qin Shaozheng's research lab, and Wu Jianhui, a professor of Shenzhen University, are the co-first authors of the paper. Other collaborators include Dr. Geng Haiyang, Prof. Liu Chao, Prof. Luo Yuejia, and Prof. Luo Jing. Corresponding author is Prof. Qin Shaozheng.
Lab website:http://icanbrainlab.bnu.edu.cn/en/infolist-99.html
Paper link:Liangying Liu, Jianhui Wu, Haiyang Geng, Chao Liu, Yuejia Luo, Jing Luo, Shaozheng Qin, Long-Term Stress and Trait Anxiety Affect Brain Network Balance in Dynamic Cognitive Computations, Cerebral Cortex, 2021;, bhab393, https://doi.org/10.1093/cercor/bhab393
Lab papers in the field of stress and affective cognition (Excerpt):
L Zhuang, J Wang, B Xiong, C Bian, P Bayley,S Qin*. Rapid neural reorganization during retrieval practice predicts subsequent long-term retention and false memory.Nature Human Behaviour (2021).
B Xiong, C Chen, Y Tian, S Zhang, C Liu, TM Evans, G Fernández, J Wu, S Qin* (2021). Brain preparedness: The proactive role of the cortisol awakening response in hippocampal-prefrontal functional interactions. Progress in Neurobiology 205, 102127.
J Wang*, B Chen, M Sha, Y Gu, H Wu, C Forcato, S Qin* (2021). Positive and neutral updating reconsolidate aversive episodic memories via different routes. Neurobiology of Learning and Memory 184, 107500.
Y Liu, W Lin, C Liu, Y Luo, J Wu*, PJ Bayley, S Qin* (2016). Memory
consolidation reconfigures neural pathways involved in the suppression of emotional memories. Nature communications 7 (1), 1-12.
Y Zeng, F Tao, Z Cui, L Wu, J Xu, W Dong, C Liu, Z Yang, S Qin* (2021). Dynamic integration and segregation of amygdala subregional functional circuits linking to physiological arousal.
NeuroImage, 118224
N Jiang, J Xu, X Li*, Y Wang, L Zhuang, S Qin* (2021).Negative parenting affects adolescent internalizing symptoms through alterations in amygdala-prefrontal circuitry: a longitudinal twin study. Biological Psychiatry 89 (6), 560-569.
T Tian, CB Young, Y Zhu, J Xu, Y He, M Chen, L Hao, M Jiang, J Qiu, X Chen, S Qin* (2021). Socioeconomic Disparities Affect Children’s Amygdala-Prefrontal Circuitry via Stress Hormone Response. Biological psychiatry 90(3),173-181.
Y Zhu, X Chen, H Zhao, M Chen, Y Tian, C Liu, ZR Han, X Lin, J Qiu, G Xue, H Shu, S Qin* (2019). Socioeconomic status disparities affect children’s anxiety and stress-sensitive cortisol awakening response through parental anxiety. Psychoneuroendocrinology 103, 96-103.

